
Stephen Okay Donovan (the Netherlands) and John WM Jagt (the Netherlands)
A fossil is a mine of details about only one specimen of 1 species and lots of such specimens symbolize extinct species. Consequently, no observations of the residing organism are doable – the whole lot we learn about that species must be gleaned from fossils.
Morphology (the shape or form of an organism or a part of it) is clearly a place to begin – that’s, what are the options of the specimen? Describing a specimen could also be laborious, nevertheless it offers a factual foundation for all later determinations and hypothesis. And it is just when a specimen’s morphology is totally understood that its evolutionary relationship to different organisms might be confidently labored out. Is it a species that’s already well-known or a uncommon specimen? Or is it a brand new species?
Astute geologists will make additional observations and deductions. If she or he collected the specimen, then it needs to be supported by data in regards to the stratigraphy, sedimentology, and associated animals and plants in the area. What do these many strands of evidence say about the environment and ecology of the specimen and species? In addition, what does the morphology of the specimen suggest about how the fossil functioned as a living organism? Many fossil organisms are comparable to living organisms, for example, interpretation of Chalk (Late Cretaceous) Micraster can benefit from what we know about living heart urchins.
Therefore, we can build up a collection of facts about our specimen’s form, classification, environment and function. The next step may be more problematic. How did the fossil interact with other organisms when it was alive? Without direct evidence, we are left only with uniformitarian arguments (that is, arguments that assume that the present is a key to understanding the past), although the older the fossil, the more speculative such an interpretation may have to be.
What makes these interpretations of ancient organic interactions more valid is those relatively rare examples in which the evidence is actually preserved. To return to the examples of Upper Cretaceous irregular echinoids (for example, the Micraster mentioned above), one line of evidence is provided by examples in which the fossil tests (a ‘test’ is the shell of the sea urchin) retain the skeletons of encrusting organisms, such as oysters, cementing brachiopods and bryozoans. Of course, care must be taken to determine if such infestation occurred before or after the death of the echinoid, but there are various lines of evidence that can be used and some are discussed below.
However, what happens if these hard parts are not retained or the infesting organism lacked any? In particular, how can the evidence of boring, biting and embedding trace fossils on an echinoid test provide evidence of an ancient ecology?
In brief, different types of trace fossil data can provide different evidence for reactions between the organisms that lived together. In this article, we will discuss some examples from our own research in northern Europe.
(1) Echinocorys from northern France
Echinoids that are perforated by small round holes or bearing pits, or both (assignable to the ichnotaxon Oichnus) are rare and enigmatic fossils. Circular drill holes that perforate the test are commonly considered to be the result of predatory or parasitic activity by certain groups of gastropods. It is more difficult to determine the function of circular pits on the surface of the echinoid test (morphologically Oichnus), but which are obviously not the result of predation.
In Fig. 1, there is a specimen of the common, Late Cretaceous, holasteroid echinoid, Echinocorys scutata, which bears two Oichnus pits that invite palaeoecological consideration. On this echinoid, there are small, shallow, circular, non-penetrative pits, which are situated in adjacent columns in an ambulacrum (that is, a radially arranged band, together with its underlying structure, through which the double rows of tube feet (tentacle-like extensions of the water vascular system) protrude).
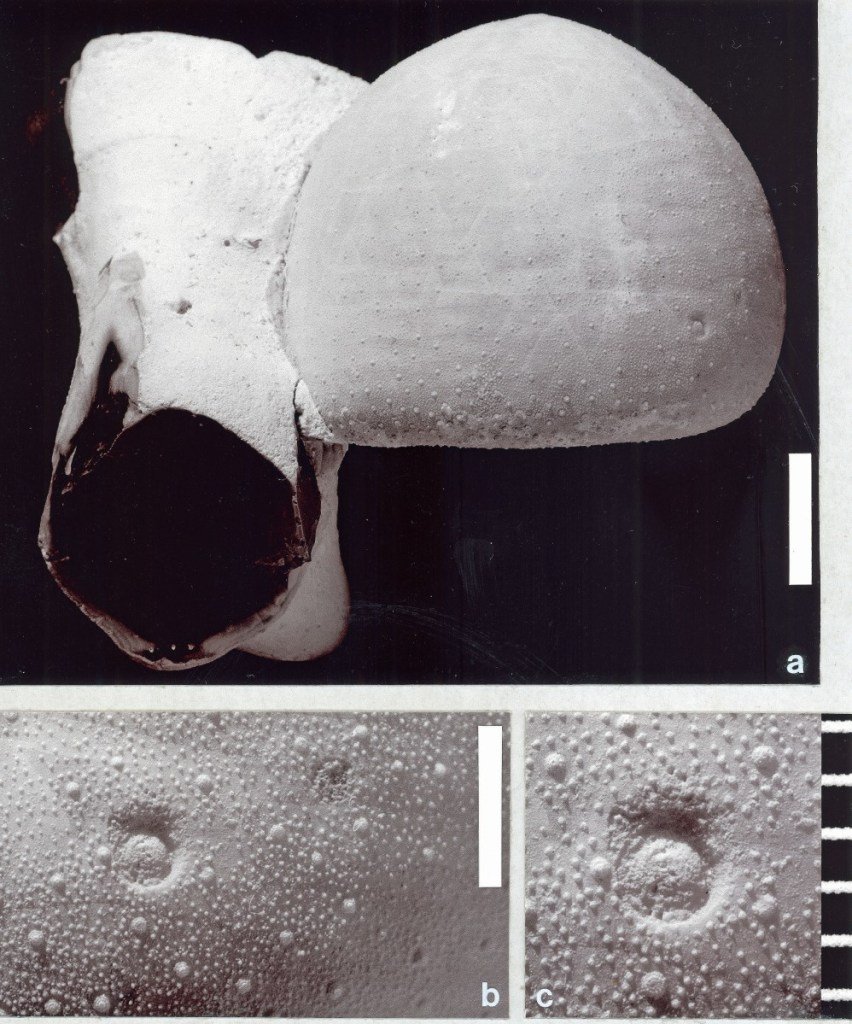
People who research drill holes or pits are divided into two groups – those that name their small round holes or pits and those that do not! We agree with the fundamental idea of systematic ichnology (that is, the study of trace fossils – see, for example, the article by Henk Oosterink in this issue). This is succinctly defined by Pickerill (1994, p. 15), such that:
… the labelling of ichnotaxa provides a necessary vocabulary for writing and conversing about trace fossils.”
Therefore, it is important to this article to assign our round pits to the appropriate ichnotaxon (that is, a taxon – or a unit of one or more organisms – based on the fossilised work of an organism, for example, a burrow, boring or footprint).
The circular pits described above are included in Oichnus excavatus (Donovan and Jagt), but only with some hesitation. This particular trace fossil is usually deeper and more prominent. However, the main interest in these holes rests in the palaeoecology of their producers. Small round holes in the tests of fossil echinoids present problems of interpretation, the most obvious questions being who did it and why?
The specimen of E. scutata shown in Fig. 1 provides an example of distinctive behaviour of such pit-forming organisms. The two shallow, non-penetrative pits are close together and each is precisely located. Such a precise location strongly suggests that the echinoid was alive when it was infested by the pit-forming organisms, although there is no indication (such as obvious deformity of the growth of the test) to support this.
The pits may represent examples of predation that has been disturbed or, more probably, could represent the traces of one or two organisms that hitched a ride on the test for protection, or to gain a feeding or respiratory advantage. In this respect, their position may indicate a preference by the pit-forming organism for the front end of the echinoid. The pit-forming organism was probably filter feeding and, by attaching itself in this position, automatically gained elevation and the correct orientation for feeding, as well as the protection provided by the surrounding tube feet and spines. However, the identity of the pit-forming organism remains unknown.
(2) Punctured Hemipneustes
The holasteroid echinoid, Hemipneustes striatoradiatus (Leske), is a large and striking part of the invertebrate macrofauna of the type Maastrichtian (Upper Cretaceous) of Limburg and Liège in the Netherlands and Belgium. Live and dead tests of this taxon provided hard substrates for infestation by a variety of encrusting and pit-forming organisms.
Evidence of the interactions of organisms provided by H. striatoradiatus substrates includes borings, encrustation by shelly invertebrates and marks of predation. However, while the unusual specimen discussed below (see Fig. 2) is distinctive, it is uncertain if it is caused by a large invertebrate or is an example of a deep puncture made by a predatory marine vertebrate and subsequently repaired by the echinoid. In fact, the pit has an unusual morphology, unlike any other trace fossil found in these common echinoids.
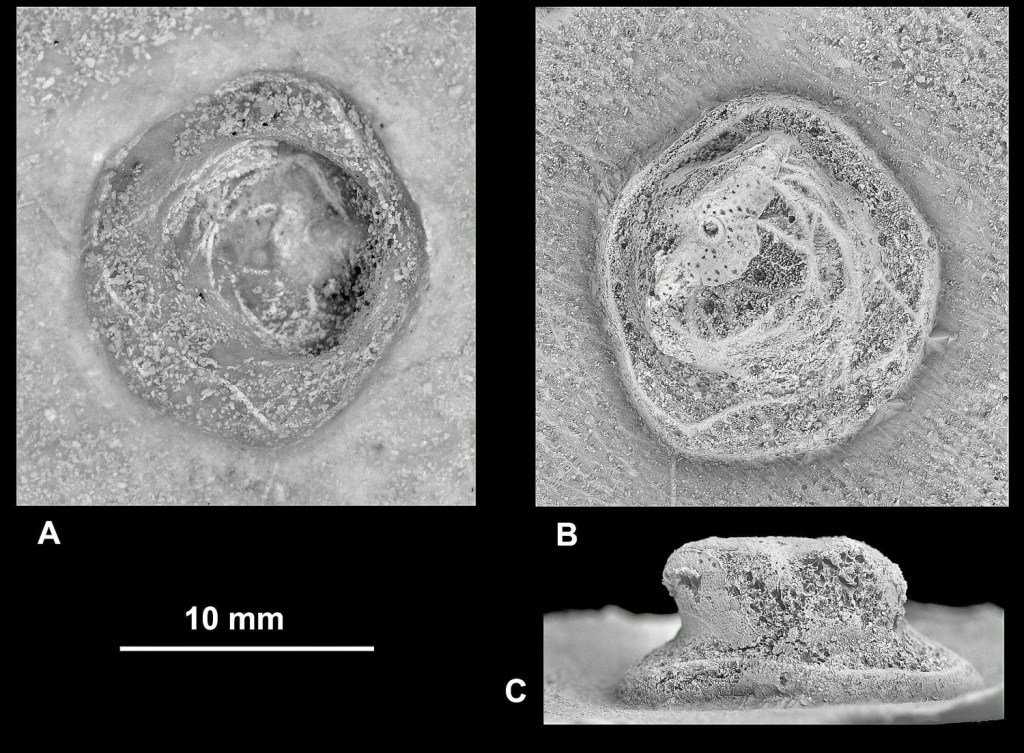
The test of this H. striatoradiatus is well preserved apart from a few cracks probably arising after deposition. The size, position and morphology of the pit on the centre of the oral (lower) surface, lying on the midline between the mouth and anus are all unusual (Fig. 2). The pit is lined with small tubercles (that is, the bases of spines), demonstrating that, whatever sort of infestation it represents, it was not lethal, because the echinoid was able to repair itself.
The pit is large, close in size to Oichnus excavatus, but it is not quite the same and is situated on the ‘wrong’ part of the test. With only rare exceptions, O. excavatus is found on the aboral (upper) surface and such a well-developed specimen is unknown from such a central position on the adoral surface (that is, the lower surface, in contrast with the sediment surface in life and bearing the mouth). The pit is also more deeply impressed than an attachment scar of, for example, a foraminiferan living on the sea floor. Intuitively, the position of the pit seems to have been a poor one for a large, filter-feeding, invertebrate-infesting organism that was ‘hitching a ride’ on a live, furrowing, echinoid living on the sea floor. (Compare this with the previous example.)
The trace described above is a deep indentation on an area of the test that commonly was not available for infestation by invertebrates during the life of the echinoid. The position and morphology of this trace lead us to think that it may have been caused by a vertebrate, rather than an invertebrate, and possibly represents a scar of a less-than-lethal predatory attack. That is, the wound healed after the attack and the evidence was preserved as a pit. This is supported by it being of similar diameter to other traces in the same species that we also think were produced by vertebrates.
Fossil vertebrates known from the type-area of the Maastrichtian included mosasaurs, elasmosaurid plesiosaurs, turtles, a crocodile, dinosaurs and various fishes. The pit could have been made by a vertebrate with rounded, well-separated teeth, because this is the only penetrative wound in the test. The distance from the circumference of the perforation to the closest parts of the external edge of the echinoid is about 30mm.
The tooth may have been pointed or blunt, evidence for the former being disguised by the echinoid, which seem to have sealed the base by growing new test over it. The two, teardrop-shaped indentations on the aboral surface may have been produced by two teeth of the other jaw failing to gain purchase and slipping across the surface. Based on these observations and deductions, the most probable culprits were a mosasaur or a bony fish – the tooth was conical, but may have been truncated, perhaps due to having been broken.
(3) A five-sided attachment scar
Tests of large Late Cretaceous holasteroid echinoid genera, such as Echinocorys Leske and Hemipneustes L. Agassiz, do not usually preserve growth reactions to attacks from settling, cementing or boring invertebrates, or lesions caused by vertebrates, which occurred during the life of the echinoid. However, there may be subtle evidence that such infestations occurred during life. Infestations not associated with modifications of the test are interpreted as having occurred, most probably, after death, but before the final burial of the echinoid test.
The trace fossil shown in Fig. 3 is from a test of Echinocorys gr. conoidea (Goldfuss). Although the test is slightly fractured and possibly lightly crushed, this has not altered the morphology of the large trace fossil, which is non-penetrative. There is also no evidence of the echinoid reacting to this infestation with any unusual form of growth.
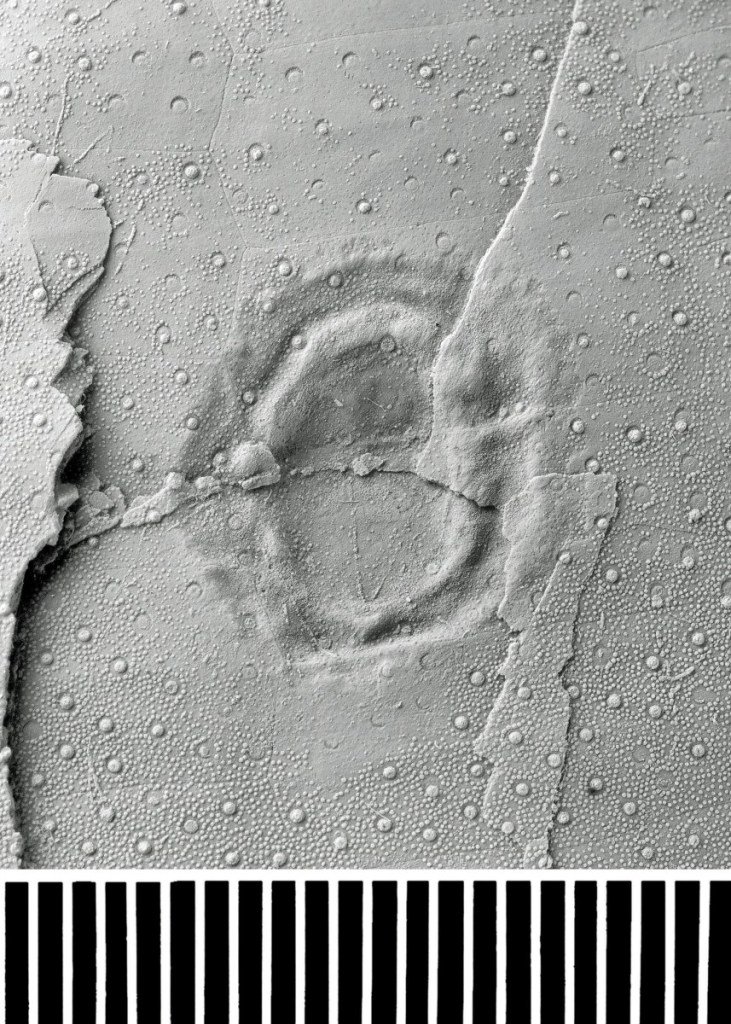
This trace fossil is unlikely to be a healed puncture, on account of the complex morphology of the margins, with the folded areas inside the circumference. We think it is the result of an unmineralized, sessile (that is, non-mobile) invertebrate embedding itself (only to a shallow extent) into the echinoid while it was still alive. This idea is supported by the presence of numerous tubercles That is, attachment points for spines) within the area of the trace, suggesting that spines probably regenerated after the death of the trace maker. The position of the trace close to the centre of the widest part of adjoining ambulacrum would have placed the maker near the maximum distance possible from any tube feet, but above the sediment surface. Bald areas of test around the trace may be the result of cleaning by the trace maker.
We prefer the idea that this trace is a scar produced by something like a sea anemone. The shape of the trace might have been normal for the producing species, but it may also be due to a response to its environment. For example, the producing organism may be elongated by gravity, having attached itself to an overly steep surface. Alternatively, if the organism was harvesting plankton from currents flowing past the test, then a more elongated form may have been a successful feeding strategy.
Acknowledgements
We are indebted to Phil Crabb (Photographic Unit, The Natural History Museum, London), who provided all of the images for this article.
References
Donovan, S.K. & Jagt, J.W.M. 2005. Site selectivity of pits in the Chalk (Upper Cretaceous) echinoid Echinocorys Leske from France. Bulletin of the Mizunami Fossil Museum 31 (for 2004): 21-24.
Donovan, S.K., Jagt, J.W.M. & Dols, P.P.M.A. 2010 (in press). Ichnology of Late Cretaceous echinoids from the Maastrichtian type area (The Netherlands, Belgium) – 2. A pentagonal attachment scar on Echinocorys gr. conoidea (Goldfuss). Bulletin of the Mizunami Fossil Museum 36.
Donovan, S.K., Jagt, J.W.M. & Lewis, D.N. 2008. Ichnology of Late Cretaceous echinoids from the Maastrichtian type area (The Netherlands, Belgium) – 1. A healed puncture wound in Hemipneustes striatoradiatus (Leske). Bulletin of the Mizunami Fossil Museum 34: 73-76.
Pickerill, R.K. 1994. Nomenclature and taxonomy of invertebrate trace fossils. In: Donovan, S.K. (ed.) The Palaeobiology of Trace Fossils, 3-42. John Wiley and Sons, Chichester.
Trending Products









